- COMPORTEMENT ANIMAL
- COMPORTEMENT ANIMALSi, d’une manière triviale, on peut dire que le comportement d’un animal c’est ce qu’il fait, la fugacité des événements et leur caractère exclusivement phénoménal exigent un effort de définition plus approfondi que pour un objet scientifique traitant de structures concrètes. C’est donc à cette première exigence qu’il faut répondre. De même, si le comportement constitue un ensemble de phénomènes d’observation banale, accessibles à quiconque, sans instrumentation particulière, la science qui l’étudie a dû inventer des méthodes de mesure originales, qui se sont complexifiées avec les exigences des questions posées. Même si elle reste dans la grande tradition naturaliste, elle ne se contente plus d’une simple description qualitative, mais elle fait l’objet d’une quantification complexe, et utilise des méthodes statistiques élaborées et une instrumentation informatique lourde.Tinbergen, l’un des trois Prix Nobel de médecine qui ont honoré la discipline en 1973, a défini les objectifs de l’éthologie par quatre questions auxquelles elle a pour vocation de répondre et qui délimitent le champ de la discipline: 1. Quelles sont les causes (immédiates) du comportement? 2. Quelle est sa valeur de survie? 3. Comment s’est-il mis en place au cours de l’ontogenèse? 4. Comment s’est-il mis en place au cours de la phylogenèse ? Chacune de ces questions correspond à un niveau d’analyse, et les divers niveaux sont emboîtés les uns dans les autres, la phylogenèse englobant les ontogenèses, les causes immédiates ne représentant qu’un instant du processus de développement, les fonctions résultantes du comportement (sa valeur de survie) venant en retour sur chacun des trois niveaux. Même si ce n’est pas la seule manière possible, présenter l’étude du comportement animal, l’organiser autour de ces quatre questions reste certainement la plus didactique.La mise en place phylogénétique du comportement doit être envisagée tout d’abord comme le résultat des processus sélectifs intervenus au cours de l’évolution. Il participe à la valeur adaptative globale de l’ensemble du phénotype en réalisant toute une série de fonctions qui contribuent finalement à la survie des individus, donc au succès des populations et des espèces. Les réponses à la deuxième et à la quatrième question de Tinbergen sont donc intimement liées dans une approche particulière de l’éthologie qualifiée de fonctionnelle. Le point de vue et les données véhiculées par ce courant de pensée, actuellement dominant, ont donc été largement développés ici, à travers les deux grandes avancées théoriques auxquelles ils ont donné naissance: la sociobiologie et l’écologie comportementale.Mais il ne suffit pas d’aborder d’une manière univoque les relations entre phylogenèse et comportement animal, car ce dernier apparaît aussi, de plus en plus, comme un acteur de l’évolution des espèces. L’ontogenèse et la phylogenèse sont intimement liées, même si notre ignorance reste grande dans ce domaine qui constitue l’un des points chauds de la discipline. L’analyse critique de l’interprétation néo-darwinienne, à l’origine de la sociobiologie et de l’écologie comportementale, a conduit à des propositions théoriques alternatives qui sont actuellement à la recherche de leurs fondements factuels et expérimentaux.Même si elle recouvre le premier grand débat de l’éthologie, aujourd’hui largement dépassé, la querelle de «l’inné et de l’acquis», la causalité ontogénétique, dans cette perspective, mérite d’être abordée avec un regard nouveau. Elle doit en effet rassembler d’abord tous les travaux qui portent sur les relations entre facteurs génétiques et épigénétiques dans la genèse des phénotypes comportementaux et des structures nerveuses qui les sous-tendent. Les données récentes dans le domaine de la génétique comportementale, c’est-à-dire des relations entre les gènes, la construction du système nerveux et le comportement, fournissent des arguments critiques et décisifs dans notre vision de l’ontogenèse et de la coaction entre maturation et expérience, entre génétique et épigénétique. Même s’il est aisé de démontrer des influences génétiques sur la genèse du système nerveux et les traits de comportement qui en émergent, seule une vision naïve tend à considérer qu’un gène code pour un caractère et qu’un trait de comportement aurait alors son image génétique traduite ensuite en phénotype. De même, l’idée qu’il existe un segment d’ADN qui construit le système nerveux est, elle aussi, probablement fausse ou simpliste. Il n’existe aucune correspondance 1/1 entre gènes et structures nerveuses, mais plutôt des relations multiples qui, à chaque stade du développement, sont guidées et actualisées par des informations épigénétiques. On comprend alors que toute dissociation entre facteurs génétiques et épigénétiques dans les émergences comportementales apparaît comme une approximation purement statistique, dont la validité reste très illusoire au niveau des individus exprimant leur phénotype comportemental singulier.Les études descriptives et expérimentales d’ontogenèses comportementales, associées avec les données de plus en plus riches de la psychologie (animale, comparative, expérimentale) qui ont été largement intégrées dans l’éthologie au cours des années soixante, nous conduisent à considérer le phénotype comportemental non seulement comme un ensemble de traits plastiques, mais comme l’instrument essentiel de cette plasticité et de la différenciation des individus. L’ontogenèse comportementale apparaît alors comme une aventure individuelle, l’individuation, imprévisible dans ses aspects les plus spécifiques, obéissant à des lois générales homologues de celles de la biologie du développement.La causalité immédiate du comportement tient compte de toutes les conditions environnementales locales et des compétences de l’individu à l’instant correspondant de l’ontogenèse. Au-delà de tous les supports de l’action ou de la réaction dont il dispose à un instant donné, l’animal est en mesure de réaliser de nouveaux comportements dont les divers aspects seront ensuite intégrés, en de nouvelles compétences, à son expérience individuelle.Enfin, au-delà des réponses aux quatre questions de Tinbergen, il a été fait dans le présent article un choix philosophique capital qui organise l’ensemble du discours, parce qu’il nous a paru plus conforme à la réalité des phénomènes. Nous avons refusé de considérer les êtres vivants comme des victimes innocentes d’une histoire qui leur aurait été imposée de l’extérieur, par quelque surdéterminisme qui échapperait à l’investigation scientifique. Au contraire, nous avons traité le vivant dans son ensemble comme autonome, c’est-à-dire capable de s’auto-organiser, à partir de propriétés qui lui sont propres. Cette perspective systémique du vivant autonome et auto-organisateur sous-tend chacun des chapitres, pour rendre compte des divers niveaux de causalité emboîtés, phylogénétique, ontogénétique et immédiate, pour comprendre également les fonctions du comportement. Elle constitue donc le cadre théorique qui charpente l’ensemble du texte.1. Les jalons de l’étude scientifique du comportement animalCourants conceptuels et définitionsL’une des premières définitions, très mécaniste, est celle du manifeste behavioriste (behavior = comportement) de Watson (1913): «l’ensemble des réactions adaptatives objectivement observables qu’un organisme, généralement pourvu d’un système nerveux, exécute en riposte aux stimulus (eux aussi objectivement observables) provenant du milieu dans lequel il vit». Le behaviorisme constitue une science du couple stimulus-réponse (S-R), lié par une boîte noire (black box ) dans laquelle il est objectivement impossible de pénétrer. Le comportement serait donc la manifestation motrice observable d’une réflexologie globale qui régirait le fonctionnement d’une «machine animale».D’autres définitions sont exclusivement opérationnelles, comme celle de Tolman (1932): «... l’activité apparente extérieure d’un être vivant considéré comme un tout» ou bien «... l’ensemble des manifestations motrices observables d’un individu à un moment et dans un lieu particuliers» (Campan, 1986).Enfin, un troisième type de définition relève d’une conception relationnelle du comportement, «expression de la relation d’un sujet à son monde». Von Uexküll (1934) le décrit sous la forme de cycles fonctionnels d’interactions réciproques entre un animal et son monde subjectif: l’univers propre (Umwelt ). Cette vision reste largement inspirée du behaviorisme, mais elle innove en introduisant la dimension relationnelle, puis en séparant l’univers propre de l’animal de celui de l’observateur. Von Uexküll distinguait un monde perceptif et un monde actif: l’action du milieu sur le comportement s’exercerait à travers les systèmes neurosensoriels de l’animal, qui réagirait à son tour par son système d’actions sur le lieu.Cependant, à l’origine, l’Umwelt caractérisait une espèce animale, alors que les mondes actif et sensible, même s’ils sont limités par un cadre relativement «spécifiques de l’espèce», appartiennent en réalité à un individu, prenant en compte la combinaison des événements maturatifs et de «l’expérience» vécus par le sujet.L’Umwelt est donc modifiable parce qu’il peut changer objectivement avec le temps, que les capacités sensorielles et motrices varient avec l’âge, mais aussi en raison des caractères que chaque individu lui attribue et des représentations qu’il s’en construit, selon les effets renforçateurs vécus aux divers instants de son existence (expérience), ainsi que l’efficacité de l’exercice moteur accompli. La dynamique du cyle fonctionnel ne peut pas se limiter à une boucle qui se referme sur elle-même; elle est aussi diachronique, traduisant, plus qu’une simple interaction instantanée, l’histoire ou le déroulement d’une relation subjective qui fait échapper le comportement à une vision exclusivement behavioriste.L’individu et son univers propre constituent donc un système actif, dynamique et relativement autonome, d’où émergent des formes comportementales qui dépendent aussi bien de son «histoire» (phylogénétique, génétique et ontogénétique), que des conditions actuelles (ici et maintenant), l’exécution du comportement contribuant à son tour à l’histoire (Campan et al., 1987). Le comportement relève donc du champ des phénomènes et non des objets physico-chimiques, même si c’est le système nerveux qui supporte l’émergence.Méthodes d’étudeLa description du comportement représente une difficulté méthodologique particulière, car elle exige de passer d’un discours qualitatif, qui rend compte spontanément de la continuité de ce que fait un animal d’une manière que l’on pourrait qualifier d’«analogique», à une traduction quantitative de type «digital».La «digitalisation» du comportement va consister tout d’abord, pour l’observateur, en l’élaboration d’un répertoire de formes motrices ou «praxèmes», adapté à la finesse qu’il veut donner à son observation. Il doit pouvoir dénommer, sous un terme identique, des unités comportementales parfois un peu différentes, mais identifiables comme appartenant à une même catégorie. Il peut alors enregistrer des éthogrammes qui représentent la succession des praxèmes observés, en respectant les durées dans le cas d’activités qui se prolongent.Quel que soit le moyen utilisé – observation directe et saisie manuelle, sur papier ou sur ordinateur, ou bien transcription à partir d’un enregistrement au magnétophone ou au magnétoscope, ou bien par l’intermédiaire de dispositifs de radiopistage et de capteurs bio-télémétriques –, le saut du qualitatif au quantitatif est réalisé par la mesure de trois types de paramètres descriptifs: la fréquence et la durée des praxèmes enregistrés, puis les fréquences des transitions qui les associent deux à deux, ou les enchaînent en séquences. La figure 1 est une illustration de la description quantitative d’un comportement constitué d’un répertoire de praxèmes organisé en séquence, dont on a évalué en terme de probabilité les fréquences de transition entre les unités.De plus, le comportement se déroule dans un espace particulier. L’observateur doit donc aussi quantifier les corrélations entre la densité et la fréquence des activités réalisées, les paramètres des trajets effectués et la structure de l’espace.Les expressions motrices ainsi mesurées peuvent, ensuite, être utilisées comme des variables dépendantes dans une approche expérimentale du comportement évaluant les effets de variables indépendantes ou de situation. Dans les conditions naturelles, alors que toute action sur les paramètres de l’environnement est limitée, la prise en compte de leurs variations «spontanées», ou la comparaison de situations différentes, place les variables comportementales en position de quasi-dépendance par rapport aux conditions locales, réalisant ainsi une pseudo-expérimentation.Enfin, toutes les manifestations comportementales, spontanées ou provoquées, subissent les effets d’une constellation de facteurs plus ou moins interdépendants; leur prise en compte exige des traitements appropriés associant analyses multivariées et statistiques causales, débouchant, de plus en plus souvent, sur des modélisations qui peuvent être améliorées après une ou plusieurs mises à l’épreuve expérimentales successives.2. Comportement et phylogenèseIl s’agit de la dernière des quatre questions posées par Tinbergen en 1963 pour définir les objectifs de l’éthologie «étude biologique du comportement»: il est à la fois un résultat de la phylogenèse et l’un des processus qui y participent.Le comportement, résultat de la phylogenèseLa comparaison globale du comportement d’espèces situées à divers niveaux de la phylogenèse fait apparaître une relation entre complexification des structures du système nerveux et des manifestations comportementales. L’accroissement global des possibilités de traitement de l’information, par des systèmes nerveux, des analyseurs sensoriels et des acteurs moteurs de plus en plus performants, enrichit les possibilités pour les animaux d’exprimer, par le comportement, leur relation au monde. L’élévation du niveau phylétique s’accompagne globalement d’une réduction progressive des comportements non appris, stéréotypés et automatiques, associés à des stimulations sensorielles élémentaires, au profit d’activités de plus en plus variables, impliquant des processus perceptifs, d’acquisition, de mémoire, mais aussi de représentation et de raisonnement de complexité croissante.La mise en perspective évolutive du comportement le fait généralement considérer comme un trait phénotypique, au même titre que les caractères morphologiques. Dans le cadre d’une définition relationnelle du comportement telle que nous l’avons proposée, ce sont évidemment les supports neuro-sensori-moteurs, à l’origine des manifestations motrices (les programmes comportementaux), qui résultent du processus évolutif, et non l’expression phénoménale immédiate et locale de la relation d’un sujet au monde.Selon la théorie synthétique de l’évolution, ces programmes comportementaux ont été l’objet des deux processus successifs: tout d’abord, une variation, par mutation, recombinaison ou dérive génétique, puis une transmission différentielle de ces variants génétiques au fil des générations. C’est lors de cette seconde étape que la sélection naturelle joue son rôle [cf. ÉVOLUTION].La sélection naturelle, c’est en effet la transmission différentielle de variants génétiques alternatifs (Ayala et Valentine, 1979). Certains d’entre eux accroissent les chances de survie et de reproduction de leurs porteurs par rapport à d’autres. La sélection naturelle agit en éliminant des individus avant qu’ils ne se reproduisent, mais aussi par ceux qui produisent le plus de descendants. Elle s’exprime, finalement, sous la forme d’une reproduction différentielle. La fitness , valeur sélective ou adaptative, est une mesure globale de l’efficacité reproductrice d’un génotype.Toutefois, si ce sont les gènes (et non les traits) qui sont transmis d’une génération à la suivante, ce sont les traits (les phénotypes et non les gènes) qui sont les cibles des agents de sélection (Waddington, 1975). On doit distinguer, puis corréler fitness et adaptation: une meilleure fitness doit conférer à son porteur des adaptations utiles, et réciproquement.Les caractères phénotypiques sélectionnés sont des programmes nerveux de comportement, comme l’illustrent divers exemples de radiation évolutive dans le domaine éthologique. La phylogenèse des pinsons des Galapagos (Darwin) ou celle des anatidés (Lorenz) constituent des modèles fondateurs. Nous citerons des travaux plus récents sur les Drosophiles hawaïennes (Hoy, 1990).Pour une ou deux espèces fondatrices, il en existe aujourd’hui dans l’archipel, de huit cents à mille, soit le quart des espèces de Drosophiles existant au monde. Pour certaines d’entre elles, des filiations phylogénétiques fondées sur une analyse de marqueurs moléculaires ont été corrélées avec la structure des divers chants de cour des mâles (séquences de vibrations alaires et de mouvements abdominaux) [fig. 2].Le mode ancestral de production sonore est la vibration alaire, générée par des muscles thoraciques primitivement associés au vol. Bien que D. melanogaster émette des motifs brefs de syllabes sonores, alors que D. sylvestris produit des trains plus longs, durant plusieurs secondes, le chant de D. sylvestris est assez proche de la forme continentale de D. melanogaster . Mais les chants ne sont pas produits par les mêmes structures: chez D. melanogaster , ce sont les vibrations alaires seules, alors que chez D. sylvestris il s’agit de mouvements abdominaux. D. sylvestris produit également un second type de chant, au moment où il va copuler. Le mâle se place derrière la femelle, enfonce sa tête sous les ailes, étend ses propres ailes dont il bat vigoureusement. Acoustiquement, ce signal ressemble à une sinusoïde; il n’existe pas chez D. melanogaster , mais se retrouve, avec le comportement associé (tête sous les ailes), chez plusieurs Drosophiles hawaïennes à ailes ornementées appartenant au même ensemble géographique et génétique restreint. D. cyrtoloma ne chante que pendant la phase ultime de la cour, lorsque le mâle a la tête sous les ailes de la femelle, ce qui correspond au chant sinusoïdal de D. sylvestris , mais ici avec un fort rythme pulsatile. Enfin, chez D. fasciculisetae , le chant est produit lorsque le mâle a la tête sous les ailes de la femelle; il consiste en une série de cliquetis très doux, accompagnés du relèvement des ailes, qui sont ensuite repliées en position de repos, en synchronisme avec des coups de tibia sur l’abdomen de la partenaire. Chacune de ces modalités correspond à un programme nerveux spécifique.Toutefois, les résultats de la sélection dans le domaine comportemental ne sont pas des commandes de traits isolés conservés pour eux-mêmes, mais des ensembles constituant des «stratégies»; chacune est un compromis entre plusieurs traits (Manning, 1967), qui, séparément, n’atteignent pas la meilleure fitness, mais dont la combinaison est optimale. Comme l’écrit Schlichting (1989), «le conflit entre solutions optimaliste et réaliste pose la question de ce qui est possible à un génotype. Il existe une limite supérieure au nombre de stratégies discrètes qui peuvent être évoquées en réponse à une combinaison de variables environnementales. Dans un nouvel environnement, l’expression des gènes doit être chaque fois ajustée pour de multiples traits à la fois, maintenant l’intégration phénotypique aussi proche que possible de l’optimum».C’est sur cette vision optimaliste des stratégies comportementales qu’est fondée l’approche fonctionnelle du comportement (Wilson, 1975, pour la sociobiologie ; Krebs et Davies, 1978, pour l’écologie comportementale, behavioral ecology ). Elle répond précisément à la deuxième des quatre questions de Tinbergen (1963): quelle est la valeur de survie d’un comportement? Elle associe directement la causalité phylogénétique avec son résultat fonctionnel, la fitness, l’une expliquant l’autre et réciproquement.L’approche fonctionnelle du comportementLes principes fondamentaux sont les suivants:– les gènes sont potentiellement immortels: ils se répliquent individuellement alors que le génotype qui les combine et la «machine à survie» qui les porte (le phénotype) disparaissent sans être copiés;– la sélection naturelle conduit à des «stratégies évolutionnairement stables» ou S.E.S. (Maynard-Smith, 1978), à des «politiques programmées», des séquences de décisions dont le phénotype est l’exécutant;– les stratégies comportementales exécutées par une «machine à survie» répondent à la règle de l’optimalité, au meilleur rapport possible entre bénéfice et coût. S’il n’en était pas ainsi, les individus, qui expriment une variante comportementale accroissant leurs chances relatives de survie et de reproduction, produiraient plus de descendants, éliminant progressivement la stratégie initiale de la population.Un exemple de stratégie de capture de proies ou de récolte d’aliments (optimal foraging ) illustrera cette approche. L’hypothèse est que «les prédateurs choisissent les proies les plus profitables»; chacune est caractérisée par un «coût», exprimé par le temps passé à la maîtrîser et à la consommer, et un «bénéfice» en termes de «valeur alimentaire nette» (valeur totale moins le coût énergétique de la consommation et de la digestion). La valeur alimentaire nette divisée par le temps de maîtrise et d’ingestion mesure la «profitabilité» de la proie. Le prédateur optimal sélectionne les plus profitables des proies qui lui sont offertes.La figure 3 montre un exemple expérimental, où cela est vérifié chez la bergeronnette. Le choix modal des proies de 7 millimètres ne correspond pas à leur disponibilité, mais à leur profitabilité. Les décisions de l’animal se font selon les mêmes règles pour le choix: du lieu de chasse, du type de proies, du moment où il faut changer de lieu, etc.De nombreux travaux ont démontré l’optimalité des stratégies stables, dans bien d’autres domaines fonctionnels du comportement, depuis les modes d’exploitation de l’espace et des ressources, jusqu’au choix du partenaire sexuel, aux comportements sociaux anti-prédateurs, etc. Ces recherches ont une particularité méthodologique commune: elles consistent tout d’abord à formuler une hypothèse quantitative, un modèle de l’expression optimale du comportement, puis à le mettre à l’épreuve de l’expérience, enfin à l’ajuster jusqu’à ce que l’observé s’accorde à la prédiction.Dans certains cas, l’avantage fonctionnel n’est pas évident. Ainsi, le dimorphisme sexuel, qui marque le plus souvent les mâles, peut être un désavantage, car il les rend plus voyants pour le prédateur. La sélection naturelle n’aurait pas dû maintenir des caractères aussi désavantageux. Mais cela est compensé par l’avantage que confèrent ces attributs lors de la compétition sexuelle entre mâles (sélection sexuelle). En réalité, du point de vue de l’approche fonctionnelle, chaque sexe n’investit pas pour le bien de l’espèce, mais pour son propre compte: un couple est une entité plus conflictuelle qu’harmonieuse. Il s’agit, pour chacun, de choisir le meilleur partenaire pour optimiser le succès reproducteur. Le mâle tend à prendre des risques dans la cour et la parade, et la femelle investit toute son énergie dans les soins et l’élevage de la progéniture.Les comportements que l’on peut qualifier métaphoriquement d’«altruistes», qui bénéficient aux autres individus plutôt qu’à celui qui les prodigue, semblent également des paradoxes de la sélection naturelle. Si l’altruisme a une base génétique, le développement d’actes «égoïstes» devrait l’éliminer. Les théories de la «sélection de parentèle» et de la «sélection de groupe» proposent des explications.La sélection de parentèle est une appellation particulière de la sélection naturelle, qui prend en considération les parents et affiliés. Puisque la sélection favorise la multiplication des gènes qui accroissent le succès reproducteur de leurs porteurs, chez un parent qui partage la moitié de ses gènes avec sa progéniture, un gène qui contrôlerait l’altruisme parental sera favorisé par la sélection, dès que la mort du parent peut sauver la vie de plus de deux de ses descendants; ce gène peut alors s’accroître dans la population plus rapidement qu’un autre seulement égoïste. Cela justifierait l’investissement parental ou l’altruisme réciproque au sein des groupes affiliatifs.Les Insectes sociaux posent, à cet égard, un problème particulier. Dans L’Origine des espèces , Darwin avait déjà souligné le paradoxe de l’existence d’ouvrières de fourmis ou d’abeilles stériles, présentant des activités altruistes telles que soigner les larves ou sacrifier sa vie pour défendre la société. Cela ne pouvait se justifier qu’en considérant la société comme un superindividu. C’est la réponse à cette question qu’apporte la théorie de la sélection de parentèle. Hamilton (1964) propose de remplacer, dans le processus évolutif, la valeur sélective d’un individu, exprimée par le nombre de ses descendants (fitness), par la valeur sélective globale du groupe social (inclusive fitness ). Ainsi, un gène favorisant la reproduction d’individus apparentés améliorerait sa propre représentation, même s’il n’est pas directement répliqué par l’individu qui le porte. Cela expliquerait qu’un trait de comportement altruiste ait été sélectionné, même s’il diminue la probabilité de survie du sujet vecteur, dans la mesure où il accroît celle de parents porteurs du même génotype (kin selection = sélection de parentèle). Ce type d’argument contribue à fonder la théorie socio-biologique (Wilson, 1975) [cf. SOCIÉTÉS ANIMALES].Dans certaines sociétés d’Insectes, le degré d’altruisme, tel qu’on peut l’évaluer empiriquement, est corrélé avec les proximités génétiques (r ). Mais la théorie n’explique pas tous les faits observés dans les colonies haplo-diploïdes d’Hyménoptères. Par exemple, si les ouvrières soignent de préférence leurs sœurs (r = 3/4), elles devraient par contre orienter leur comportement altruiste vers leur éventuelle descendance (r potentiel = 1/2) plutôt que vers leurs frères (r = 1/4); ce n’est pas le cas. Trivers et Hare (1976) ont cherché à expliquer cette apparente anomalie. Pour un parent, si le coût de la production de reproducteurs mâles est égal à celui des femelles, la stratégie «optimale» doit tendre vers un sexe-ratio de 1/1 dans sa descendance. Pour une ouvrière, par contre, sœur de ces futurs reproducteurs, la stratégie optimale est la production d’un sexe-ratio proportionnel au rapport des degrés de parenté, soit 3/4 sur 1/4 = 3/1 en faveur des femelles. Mais le coût de production d’un mâle est plus élevé, dans la mesure où il est de poids supérieur. Le modèle Hamiltonien, corrigé par Trivers, est vérifié dans plusieurs cas avec un sexe-ratio équilibré à 3/1, mais en termes de poids respectif des deux sexes et pas en nombre d’individus produits. Dans les espèces où la femelle est très grosse, le sexe-ratio est déplacé en faveur des mâles.Enfin, la sélection de groupe fait référence à la compétition entre colonies, ou entre populations, plutôt qu’entre individus (Wynne-Edwards, 1962). Supposons que, dans une espèce, certaines colonies ou populations possèdent des gènes qui favorisent la reproduction bien au-delà de la capacité d’accueil de l’environnement; elles peuvent détruire leurs ressources alimentaires et disparaître. Cela ne se produira pas si elles possèdent des gènes qui limitent la croissance de la population pour l’ajuster à la capacité d’accueil; ce comportement paradoxal sera conservé.Le comportement, acteur de la phylogenèseLe comportement joue un rôle actif dans la phylogenèse en assurant des fonctions soit de novation, dans la spéciation, soit de conservation, en isolant les espèces les unes des autres.Les espèces sont des ensembles de populations naturelles endogames, isolées, du point de vue reproducteur, d’autres ensembles du même type (Mayr, 1970). La spéciation consiste d’abord en un isolement reproducteur dont les mécanismes sont multiples, souvent éthologiques.Même si la sélection naturelle réduit la diversité comportementale en ne conservant qu’un petit nombre de stratégies optimales, elle n’est pas un simple processus négatif d’élimination du moins apte. Elle génère de la nouveauté en accroissant, dans les populations, la probabilité de nouvelles combinaisons génétiques, donc le polymorphisme. Mais «... la sélection n’anticipe pas les environnements futurs, elle n’a pas de projet préconçu: elle n’est pas finaliste mais opportuniste» (Ayala et Valentine, 1979). Elle va exploiter le grand nombre de possibilités offertes par le polymorphisme, pour générer des radiations évolutives, dont nous avons vu quelques exemples comportementaux. Tous les traits éthologiques qui contribuent au maintien du polymorphisme jouent donc un rôle actif dans l’évolution.Un travail d’O’Donald et Majerus (1985) et de Majerus et al. (1986) sur la coccinelle montre comment une pression de sélection peut agir sur le polymorphisme des préférences sexuelles, première étape vers la spéciation. Les femelles à deux taches choisissent plutôt les mâles jaunes à quatre taches. Par des expériences de sélection directionnelle, on peut faire varier le choix pour la forme jaune de 20 à 65 p. 100 en neuf générations. Par ailleurs, il n’existe pas, chez ces coccinelles, de dimorphisme sexuel morphologique évident, ce qui conforte l’idée que les choix sexuels ne se contentent pas d’expliquer la sélection de caractères exagérés, mais aussi le maintien et la stabilité de systèmes polymorphes sans dimorphisme sexuel. Plusieurs mécanismes comportementaux du même type contribuent au maintien du polymorphisme, qui influe sur la sélection sexuelle. Ainsi, la «vigueur sexuelle» (évaluée par plusieurs paramètres quantitatifs, comme le temps de latence de la copulation, le nombre d’essais de monte, la durée de la copulation, etc.) est plus élevée chez les Drosophiles mâles dont le génotype est hétérozygote (Boesiger, 1962). Une réduction du degré d’hétérozygotie diminue leur vigueur et augmente la réceptivité des femelles, alors que l’hybridation des souches consanguines a l’effet inverse. La sélection sexuelle profite alors des associations entre hétérozygotie, vigueur des mâles et réceptivité des femelles, renforcées par une plus grande fécondité des femelles et des mâles hétérozygotes, pour maintenir le polymorphisme. L’avantage du type rare est un autre mécanisme éthologique de régulation du polymorphisme lié à la sélection sexuelle (Petit et Ehrman, 1969). Les mâles qui présentent un type assez abondant dans une population sont désavantagés dans les choix que font les femelles au profit de ceux qui possèdent des caractères rares. Ainsi, chez les Drosophiles, le mutant white , porteur d’un gène récessif lié au sexe qui lui donne des yeux blancs, est désavantagé lorsque la fréquence des mâles de ce type atteint et dépasse 40 p. 100 de la population. Inversement, il fait l’objet de choix préférentiels lorsqu’il représente moins de 40 p. 100 du nombre de mâles. Une sorte d’équilibre est maintenu autour de cette valeur par une succession d’oscillations qui résultent des changements de critère de la sélection sexuelle.L’hétérosis traduit l’avantage des hétérozygotes non seulement pour la vigueur sexuelle, mais également pour d’autres traits de comportement. Ce processus a été démontré expérimentalement en comparant des souches parentes et leurs hybrides, dont les performances sont très souvent meilleures, dans des tests d’apprentissage, par exemple. La dépression par consanguinité est le contraire de l’hétérosis: elle désavantage les homozygotes issus de croisements consanguins, dont les performances sexuelles et comportementales globales sont toujours inférieures.L’ensemble de ces processus, en assurant la régulation et le maintien du polymorphisme, conservent et même enrichissent les potentialités micro-évolutives des populations. Cependant, la diversification des espèces n’est réalisée que si ces possibilités sont exprimées, par dérive progressive d’une partie de la population parente, jusqu’à l’isolement sexuel total. La spéciation consiste en une sélection sexuelle de plus en plus accentuée, soit directement, lorsque la différenciation progressive porte sur des traits du comportement reproducteur, soit indirectement, s’ils isolent deux parties d’une population, dans le temps et/ou dans l’espace.L’examen de différentes lignées de Drosophila sturtevanti du Mexique, du Guatemala et du Brésil par Dobzhanski (1944) montre que, si deux types de femelles sont mis en présence d’une classe de mâle, les croisements homogamiques sont plus fréquents que les hétérogamiques. Chez D. paulistorum , les isolements sont déjà plus accentués; considérées d’abord comme des races géographiques, elles ont pris le statut d’«espèces naissantes» regroupées dans une superespèce Paulistorum (Dobzhanski et al., 1964). Les compétitions entre lignées d’origines géographiques différentes ont souvent révélé l’avantage de l’une d’elles, dans l’un des sexes: les mâles de D. prosaltans inséminent d’autant plus de femelles qu’ils sont d’origine septentrionale. Enfin, comme nous l’avons signalé dans un paragraphe précédent, on observe des passages de croisements homogamiques au type sélectif au sein d’une même population: par exemple, en mélangeant des mouches sauvage et white , les copulations sont homogamiques ou bien donnent l’avantage au type rare.Des pressions sélectives indirectes peuvent conduire à un isolement reproducteur d’abord relatif, avec réduction du flux génique, puis total. Ces phénomènes se produisent lorsque des ensembles d’individus au sein d’une population parente sont en compétition pour la nourriture, pour l’espace ou l’une quelconque des ressources. Le partage convenable du biotope conduit progressivement à des changements d’habitudes alimentaires, à des glissements du moment de l’activité au cours de la journée ou au cours de l’année, à des déplacements de caractères [cf. ÉVOLUTION]. Alexander et Bigelow (1960), par exemple, ont décrit deux espèces de grillons Acheta pennsylvanica et A. veletis , sympatriques, dont les chants d’appel sont identiques, mais dont la maturation sexuelle des adultes a lieu à des saisons distinctes. Des pressions différentielles de prédation ou de parasitisme peuvent aussi conduire à des dérives évolutives et à des spéciations dans lesquelles le comportement joue un rôle majeur.La séparation spatiale est l’un des mécanismes les plus efficaces de différenciation de pools géniques, de croisement sélectif puis d’isolement reproducteur, donc de spéciation. L’exemple le plus connu est celui des goélands. Originaires des bords de la mer Caspienne, ils se sont propagés autour de l’océan Arctique, vers l’est pour Larus fuscus et vers l’ouest pour Larus argentatus , se retrouvant finalement au bord de la Manche. Les races géographiques sont capables de se croiser de proche en proche, mais, dans la zone de sympatrie, deux espèces sont maintenant isolées non seulement par des traits morphologiques ayant valeur de signaux dans la cour, mais aussi par la séquence d’actes de la parade. Cela signifie que les supports neuro-sensori-moteurs de ces comportements sont maintenant distincts. C’est un processus du même type qui peut rendre compte de la radiation évolutive des Drosophiles hawaïennes dont nous avons déjà parlé (Carson, 1983).Les caractères de la spéciation sont souvent plus évidents dans la zone de sympatrie où les mécanismes d’isolement des espèces, qui sont d’ailleurs les mêmes que ceux qui ont été transformés par la spéciation, accusent les différences, le comportement jouant alors un rôle conservateur, après avoir été novateur. Ainsi, par exemple, pourrait-on en partie expliquer l’absence quasi complète d’hybrides naturels entre Drosophila pseudoobscura et D. persimilis , sympatriques mais isolées par leurs choix quant aux conditions d’éclairement et d’humidité.Les traits de comportement, à travers leurs supports neurobiologiques, apparaissent donc comme un produit de la sélection naturelle, et, en retour, ils sont des acteurs de l’évolution. Toutefois, comme l’écrit Lewontin (1983), si le processus évolutif implique un double passage, des phénotypes aux génotypes et inversement, le phénotype comportemental ne peut être séparé de son ontogenèse.Comportement, ontogenèse et phylogenèsePour Schlichting (1989), «l’information sur les processus de développement est maintenant incorporée dans les théories de l’évolution des caractéristiques phénotypiques [...]. Parce que la plasticité phénotypique représente des différences génétiques dans les programmes comportementaux, elle constitue un trait, sous contrôle génétique, comme les autres traits». La plasticité ontogénétique est l’un des résultats, parmi d’autres, des processus sélectifs.C’est certainement un point de vue sensiblement plus nuancé que défend Waddington (1975): «L’insertion de l’ontogenèse dans les processus évolutifs est capitale, car c’est pendant leur construction que les phénotypes peuvent être affectés par les environnements, comme ils le sont par les génotypes.» C’est donc pour prendre en compte le rôle évolutif des modifications ontogénétiques que Waddington a fondé le concept d’assimilation génétique. Plus concrètement, il a étudié expérimentalement l’adaptation de Drosophiles à un milieu d’élevage salé, à travers les changements de taille de papilles anales (mesurées sur les pupes), impliquées dans la régulation de la pression osmotique de l’hémolymphe. La variation morphologique observée – dans les limites permises par les compétences génétiques mais résultant du stress provoqué durant l’ontogenèse par le milieu salé – est purement phénotypique. L’interprétation de Waddington est que la pression de sélection favorise les génotypes individuels qui possèdent une meilleure capacité d’adaptation ontogénétique au milieu salé: le «paysage épigénétique», modifié, «canalise» la variation morphologique, qui est ensuite assimilée génétiquement au cours du processus sélectif.Vancassel (1990) a montré l’intérêt des travaux de Waddington pour l’éthologie. Il a, d’une manière convaincante, fait la démonstration qu’un tel processus peut s’appliquer à des traits de comportement, en particulier la possibilité, pour des femelles de forficule, d’engager ou non un second cycle parental, avec tout ce que cela implique de soins maternels et de mécanismes associés. Le schéma général de l’assimilation génétique proposé par Waddington est représenté par la figure 4.C’est dans le même courant de pensée que s’inscrivent les propositions de Croset (1990) d’aborder les processus évolutifs d’un point de vue systémique. Faisant référence à Prigogine et à Atlan, Croset considère les systèmes vivants comme des structures dissipatives, auto-organisatrices, qui ont un passé. La nouveauté et la complexité naissent de la rencontre d’éléments qui, en contrecoup, subissent, de la part du système auquel ils contribuent, des contraintes telles que la nouvelle combinaison disparaîtra ou sera sélectionnée et stabilisée. Dans cette perspective: l’organisation du système résulte d’un dialogue incessant entre les niveaux d’intégration; son état au temps t dépend d’une histoire actualisée; cet état prélude à son évolution future; à tout moment, dans ces conditions, peut se produire une bifurcation qui devient irréversible, une fois entérinée par assimilation génétique. À l’appui de cette perspective, Croset tire des arguments, dont beaucoup sont comportementaux, de ses travaux sur les moustiques et les souris.3. Génétique et ontogenèse comportementalesSi le comportement résulte de la phylogenèse, il doit être héréditaire, transmissible d’une génération à l’autre, donc exister au moins en partie, sous la forme de compétences génétiques. Il est généralement admis aujourd’hui qu’un génotype rassemble des informations pour construire un phénotype dans une interaction avec l’environnement au sens large, depuis le cytoplasme de l’œuf jusqu’à l’univers écologique de l’adulte. Les compétences du génotype sont actualisées par l’environnement à travers des processus épigénétiques. C’est l’histoire de cette interaction qui constitue l’ontogenèse.Le génotype est généralement unique, c’est-à-dire que «... un être sexué ne peut se reproduire. L’enfant, n’étant la reproduction de personne, est en fait une création définitivement unique. Cette unicité résulte du nombre fabuleux d’enfants qui pourraient être procréés par un même couple...» (Jacquard, 1981). L’interaction entre la combinaison originale de gènes, qui résulte de la reproduction sexuée et de la méïose, d’une part, une succession particulière d’événements épigénétiques, dont la plupart relèvent du hasard et restent imprévisibles, d’autre part, font de l’ontogenèse, en particulier de l’éthogenèse – celle du phénotype comportemental –, une aventure complètement unique, propre à chaque individu. On comprend alors que les dimensions génétiques et épigénétiques du comportement ne soient pas dissociables.Gènes et comportementUne vision naïve tendrait à considérer qu’un gène code pour un caractère. Un trait de comportement aurait alors son image génétique traduite ensuite en phénotype. Les phénomènes sont beaucoup plus complexes.Tous les traits éthologiques pour lesquels on l’a tentée ont donné prise à une sélection artificielle au sein d’une population polymorphe. Il existe des exemples classiques, comme la sélection des Drosophiles sur la géotaxie ascensionnelle et descendante (Dobzhanki et Spasski, 1962), ou bien sur les performances des rats dans une tâche d’apprentissage (Tryon, 1940). La figure 5 donne une illustration plus récente, dans le cas d’une sélection qui porte sur le degré d’agressivité chez la souris.Dans tous les cas, la divergence est rapide, mais, si l’on relâche la pression de sélection, le retour au polymorphisme initial l’est aussi. L’intervention de facteurs génétiques dans le comportement a été aussi démontrée, en comparant soit des lignées sélectionnées sur des caractères morphologiques ou physiologiques, soit des hybrides réalisés par croisements de souches déjà sélectionnées. Dans ce dernier cas, des méthodes d’hybridation ont été mises en œuvre avec un soin méthodologique particulier, chez les Vertébrés, afin d’évaluer la variance génétique additive, avec des croisements réciproques et des adoptions croisées, ou bien, chez la souris en particulier, des implantations d’ovaires, d’ovules, d’œufs ou d’embryons. Enfin, la méthode des lignées de recombinants inbred (RI) est maintenant de plus en plus utilisée. Il s’agit de souches endogames issues de couples frères-sœurs de la même génération génétiquement ségrégante F2. Elles sont appelées RI parce qu’une partie des chromosomes de leurs souches parentales s’est recombinée à la F2, d’où les lignées RI sont tirées. Si un seul locus est responsable d’un comportement qui diffère entre deux lignées parentales, la moitié des individus de la lignée RI doivent ressembler à l’un des parents et l’autre au second, sans phénotypes intermédiaires, parce que chaque lignée RI sera homozygote pour l’allèle de l’une ou l’autre des souches parentales. Aucun des comportements étudiés jusqu’ici par cette méthode ne montre d’une manière irréfutable l’effet d’un gène unique, même majeur. Enfin, beaucoup d’espoirs reposent aujourd’hui sur l’utilisation des méthodes de génétique moléculaire appliquées au comportement.On peut proposer quelques règles générales concernant les relations gènescomportements.Les traits de comportement sont toujours affectés par plusieurs gènes. Lorsqu’un seul gène semble en cause (mutants neurologiques chez la souris ou la Drosophile, par exemple), cela signifie que la mutation touche un événement clé du développement comportemental, mais pas du tout que le comportement normal est déterminé par un seul gène.Les traits de comportement sont essentiellement déterminés par des facteurs épigénétiques. Leur héritabilité estimée, dans des expériences rigoureuses où l’on a évalué convenablement la variance génétique additive, reste toujours inférieure à 50 p. 100.Les relations gènes-comportements sont toujours indirectes. Les principaux intermédiaires sont les neurones et les réseaux nerveux au sens large (neuro-sensori-moteurs).Cela a été élégamment démontré par les techniques de dissection génétique chez la Drosophile, qui produisent des sujets gynandromorphes ou mosaïques et permettent de localiser la cible des mutations dites comportementales. La figure 6 montre comment l’analyse de sujets mosaïques qui présentent la mutation cac (cacophony ) en désigne l’origine essentiellement dans les deux moitiés du thorax. De même, pour situer la cible de certains mutants cérébelleux chez la souris, des chimères ont été produites par combinaison précoce en culture de deux morulas (à 8 cellules), avec un marqueur phénotypique permettant de différencier les cellules de Purkinje normales et mutantes. Cela montre, par exemple, que la mutation staggerer s’exprime dans les cellules de Purkinje et non dans les cellules granulaires, dont la dégénérescence est un effet secondaire, alors que c’est le contraire pour le mutant weaver . La cible d’action des gènes semble être la membrane cellulaire.Gènes et développement du système nerveuxL’idée qu’il existe un segment d’ADN qui construit le système nerveux est, elle-aussi, probablement fausse ou simpliste. Il n’existe aucune correspondance 1/1 entre gènes et structures nerveuses, mais plutôt des relations multiples qui, à chaque stade du développement, sont guidées par des informations épigénétiques. Comme l’écrit Tierney (1986), «... le développement du système nerveux est un phénomène essentiellement épigénétique dans lequel, une fois mis en mouvement par la totalité du génome, les événements s’enchaînent les uns aux autres. Un système nerveux fonctionnel dépend de l’étonnante habileté des neurones en développement à sélectionner, parmi des milliers de cibles possibles, précisément le bon neurone pour établir une connexion. Mais cela résulte de la proximité de deux cellules à un moment particulier, plutôt que de gènes spécifiant la formation des circuits».Plusieurs arguments soutiennent cette position. Ainsi, si le développement des connexions nerveuses était sous un strict contrôle génétique, on devrait s’attendre à une identité entre animaux isogéniques, élevés dans des conditions environnementales uniformes; or, si le nombre de neurones et les grandes structures anatomiques sont similaires, le nombre et la localisation des contacts synaptiques entre les cellules sont variables. De même, dans les centres nerveux de Vertébrés en développement, la mort cellulaire varie de 40 à 75 p. 100. L’idée que certaines cellules sont prédestinées à mourir s’est avérée fausse. Tous les neurones d’une population envoient des axones normaux qui atteignent le champ de leurs cibles. Là, ils entrent en compétition les uns avec les autres pour des sites synaptiques, et seuls ceux qui forment des liaisons efficaces vont survivre (stabilisation synaptique sélective). Nombre d’expériences montrent que c’est l’activité de la liaison et de la cible elle-même qui fait la sélection et régule le nombre de contacts. Enfin, dernier exemple, de très élégantes expériences ont montré que des cellules qui devaient devenir normalement des neurones à épinéphrine peuvent être transformées en neurones à acétylcholine, si elles sont placées dans des environnements appropriés.Les structures nerveuses, une fois constituées, doivent être maintenues par leurs propres fonctions. Il en est ainsi pour les projections visuelles dans le cortex occipital du chaton, d’après les travaux fondateurs de Hubel et Wiesel, puis de bien d’autres après eux. Les détecteurs traitant les informations de direction des contours d’objets (verticaux, horizontaux et divers obliques), qui se développent normalement avant l’ouverture des paupières, cessent de fonctionner en quelques semaines si l’animal est maintenu en situation de privation sensorielle différentielle pour l’une ou l’autre de ces caractéristiques. La stabilisation des structures par les fonctions doit se faire pendant une période sensible du développement, particulière à chacune d’elles, au-delà de laquelle les conséquences de la privation peuvent devenir difficilement réversibles.Toutefois, la plasticité épigénétique du système nerveux se maintient durant toute l’existence, dans les limites des compétences et des règles générales de fonctionnement fixées par le génotype. Elle reste l’une des propriétés majeures de tout système nerveux, même le plus élémentaire, comme le prouvent les multiples expériences de psychologie animale comparative qui utilisent des méthodes de conditionnement et d’apprentissage. On retrouve encore l’expression de cette plasticité dans les régulations compensatrices qui doivent être réalisées en correction des altérations dues au vieillissement, période particulière de l’ontogenèse.Ontogenèse comportementaleL’étude du développement comportemental reflète le degré de liberté épigénétique que prend le système nerveux par rapport à la directivité du génome.Plusieurs études ont montré que des influences génétiques s’exercent sur le déroulement de l’ontogenèse. Elles agissent, par exemple, sur la sensibilité précoce aux hormones (propionate de testostérone) chez la souris. On a identifié des causes du même ordre chez les abeilles, où la polyandrie sépare des sous-familles génétiques d’ouvrières différemment sensibles à l’action de l’hormone juvénile (JH), qui présentent en même temps divers degrés de plasticité comportementale dans l’adaptation aux tâches collectives de la colonie (Robinson et al., 1989).Dans les travaux classiques de Scott et Fuller (1965), plusieurs races de chiens n’ont ni les mêmes dispositions ni des parcours ontogénétiques comparables pour l’établissement des comportements sociaux en relation avec l’homme. L’héritabilité d’un trait de comportement chez les Rongeurs (évasion dans un dispositif de type shuttle-box , par exemple) diffère selon le génotype, et elle augmente au cours de l’apprentissage. Elle s’accroît avec l’âge, par sommation d’un effet génétique constant, ou elle diminue en cumulant les effets de l’environnement. Mais elle se réduit, le plus souvent, avec le vieillissement, en raison de l’accumulation de traces accidentelles affectant les systèmes physiologiques et comportementaux (Hewitt, 1990). Les facteurs génétiques sont donc impliqués dans les différences interindividuelles.De l’ensemble de la très abondante littérature sur le développement comportemental, voici quelques exemples particulièrement illustratifs.Le chant chez les oiseauxLa reconnaissance du chant spécifique commence pendant la vie embryonnaire, et son développement repose sur une expérience prénatale. Elle est très sélective chez les canetons «sauvages» ou «Pékin», alors qu’elle l’est moins pour les wood-duck (Gottlieb. 1976). La réponse au cri spécifique s’exprime précocement, depuis l’embryogenèse, dès le vingt-deuxième jour, par l’accroissement du nombre de claquements de bec; le vingt-quatrième jour, par la vocalisation propre de l’embryon ; le vingt-sixième jour, par une variation du rythme cardiaque. Ce déroulement peut être avancé, par sur-stimulation sonore, ou retardé, par privation sensorielle relative. La dévocalisation temporaire de l’embryon (en couvrant le syrinx d’un film de collodion) retarde l’apparition de la discrimination. Dans tous les cas, il existe une bonne reconnaissance du cri spécifique quarante-huit heures après la naissance, mais, si aucune expérience auditive pertinente n’intervient avant soixante-huit heures, elle disparaît. Ce ne sont donc pas les aspects qualitatifs de la reconnaissance du cri spécifique qui sont affectés, mais sa vitesse de développement et son maintien. Chez la mouette rieuse, certains cris sont déjà distingués à la fin de la vie embryonnaire, d’autres sont appris très tôt, pendant l’éclosion, mais l’expérience précoce est nécessaire pour un développement normal. Chez d’autres espèces, enfin, comme le guillemot, les embryons apprennent individuellement les caractéristiques de l’appel des parents, pendant le découpage de l’œuf.Les études comparatives du chant des oiseaux révèlent d’importantes variations interspécifiques dans la taille du répertoire. Chez certaines espèces, il n’existe qu’un seul type de chant, alors que d’autres en produisent un grand nombre. Le répertoire moyen d’une fauvette des marais (Acrocephalus palustris ) comprend des imitations de deux cent douze autres espèces d’oiseaux. Les différences dans la tendance spécifique d’un âge à incorporer de nouveaux chants dans le répertoire constituent une cause majeure de la variation spécifique de la taille du répertoire. De plus, certaines espèces n’apprennent que pendant une période critique, alors que d’autres élargissent leur répertoire tout au long de la vie.Marler et ses collaborateurs ont étudié le développement du chant chez le moineau à front blanc. De jeunes mâles capturés dès l’éclosion, puis élevés seuls dans des pièces insonores, quelle que soit leur région d’origine, produisent des versions très semblables et simplifiées du chant normal. Tout se passe donc comme s’il existait une vocalisation de base de l’espèce, manifestation phénotypique d’un codage héréditaire. Par la suite, chaque individu doit apprendre un dialecte local, celui des oiseaux adultes du voisinage, qui modifie son chant. Petrinovitch et Batista (1987), sur les mâles territoriaux du moineau à front blanc, montrent que: pour deux aires distinctes, les chants sont différents. Les caractéristiques de chaque population locale varient au cours de l’année et entre les individus, mais la possession d’un territoire confère une grande stabilité temporelle; les jeunes choisissent, sans préférence particulière, d’imiter soit leur père, soit leur voisin, ou de produire un chant intermédiaire entre les deux, qui persiste dans la population au fil des générations.La figure 7 illustre le déroulement de l’ontogenèse. Un processus d’apprentissage intervient déjà pendant les trois premiers mois, avant que l’oiseau ait chanté. Les mâles capturés au cours de leur premier automne, puis élevés seuls, chantent pour la première fois une version reconnaissable du dialecte local. Isolés jusqu’à trois mois, ils peuvent être entraînés à leur propre dialecte, ou à d’autres qu’on leur fait entendre, bien que les résultats de cet entraînement ne se manifestent que lorsque les oiseaux commencent eux-mêmes à chanter, quelques mois plus tard. Par contre, après quatre mois, ils ne sont plus réceptifs à l’entraînement, et leurs vocalisations ultérieures ne semblent plus affectées par leur expérience auditive.Des oiseaux ont été complètement et définitivement assourdis, à divers âges, par ablation de la cochlée de l’oreille interne. Si cette opération est pratiquée chez un jeune à la sortie du nid, il ne chantera, par la suite, qu’une série de notes discontinues. Ces émissions sont différentes de celles, encore parfaitement reconnaissables, produites par des sujets isolés. L’oiseau doit, en effet, être capable de s’entendre pour chanter le signal hérité; il y a donc encore une interaction entre l’héritage génétique et l’expérience. Si l’opération est pratiquée sur de jeunes oiseaux déjà entraînés à un chant normal, avant d’avoir vocalisé, leurs émissions ultérieures ressemblent à celles des moineaux rendus sourds à la naissance. À nouveau, ils doivent s’entendre pour comparer leur émission avec le modèle mémorisé. Enfin, une fois qu’ils ont commencé à chanter, le fait de les rendre sourds n’a plus d’effet.Marler (1984) distingue la structure vocale qualifiée d’innée, commune à tous les individus de l’espèce, à laquelle se surajoutent des variations résultant d’un apprentissage par imitation des congénères les plus proches (dialecte local), puis d’une invention par chaque individu, qui le distingue de ses voisins, enfin d’une improvisation qui caractérise chaque émission instantanée particulière d’un même sujet.Le comportement de quémande du jeune goélandLes poussins de goélands quémandent une régurgitation de nourriture en picorant la tache rouge qui orne le dessous du bec des parents. L’analyse image par image d’enregistrements de la séquence sensori-motrice révèle qu’elle n’est pas initialement aussi stéréotypée qu’on l’avait d’abord rapporté (Hailman, 1967). Dans leurs premières tentatives, les poussins sont peu coordonnés et dirigent leurs coups de bec vers des modèles très divers. Avec de la pratique, ils apprennent à être plus précis et sélectifs; ils répondent alors à des leurres très semblables à la tête des goélands. Les jeunes élevés dans l’obscurité par l’expérimentateur ne développent pas un comportement normal. Mais, en dépit de l’importance de l’apprentissage, les poussins ont des préférences inhérentes à picorer certaines cibles simples, par exemple les formes verticales et mobiles, plutôt que des objets stationnaires. Un article récent d’Alessandro et al. (1989) reprend la question du signal déclencheur déjà largement analysé par les travaux de Tinbergen et Perdeck (1950). Des poussins de goéland argenté, élevés au laboratoire, répondent indifféremment à des leurres (têtes) de leur propre espèce et de mouette rieuse, pourtant très différente, alors que, s’ils sont alimentés naturellement par les parents, ils font la discrimination cinq à six jours après l’éclosion. Le renforcement positif du picorage du leurre hétérospécifique génère, en quelques jours, une préférence pour le modèle récompensé. Ces auteurs suggèrent, à la suite de Margolis et al. (1987), six processus différents pouvant contribuer au développement de ce comportement: maturation, habituation, conditionnement classique, inhibition conditionnée, empreinte et apprentissage par essais et erreurs. Ils soulignent toutefois qu’une gamme particulière d’objets seulement possède le signal inconditionnel nécessaire à l’apprentissage et que l’inhibition conditionnelle peut être une force puissante de construction du comportement renforçant la spécificité.Finalement, pour Alessandro et al. (1989), le développement du comportement de quémande repose sur les mêmes principes que ceux proposés par Marler pour le chant des oiseaux, avec un modèle de base qui se précise au cours de l’ontogenèse pour conduire à une reconnaissance spécifique de l’espèce, puis individuelle.Tinbergen pensait que le comportement de quémande mettait en jeu un Innate Releasing Mechanism (I.R.M.) et un Fixed Action Pattern (F.A.P.), par lesquels les poussins possédaient une information héritée sur la configuration de la tête de l’adulte et une réponse innée, stéréotypée, à ce signal. Cependant, la simplicité des stimulus qui déclenchent initialement le picorage suggère plutôt que le système visuel du goéland comprend des neurones détecteurs de traits, à fonction perceptive générale, qui permettent aux jeunes d’acquérir le comportement de picorage du bec parental comme une réponse conditionnée opérante apprise (Tierney, 1986). De plus, les poussins actifs attirent probablement plus l’attention parentale, renforçant la réponse de régurgitation, donc le picorage. Les détecteurs de traits qui canalisent le comportement de façon adaptative peuvent avoir évolué par assimilation des gènes qui touchent à la perception visuelle des poussins. Le travail de Hailman met en lumière la possibilité pour de nombreux comportements de se développer sous une forme typique de l’espèce, à partir de facteurs simples innés (détecteurs de traits) et des conditions similaires auxquelles sont confrontés les animaux, plutôt que par des instructions génétiques précises et détaillées.Cette idée peut être illustrée par un dernier exemple d’ontogenèse comportementale, celle des comportements d’orientation chez les Insectes et les Crustacés.Chez le talitre, petit crustacé qui habite les plages, la direction astronomique, qui guide les migrations régulières vers la mer ou en direction opposée, est génétiquement déterminée (Scapini, 1986). Chaque population développe en élevage, en l’absence de vision du ciel ou de l’environnement naturel, donc indépendamment des effets épigénétiques normaux, une direction préférentielle pertinente par rapport à l’orientation de la plage d’origine. Les descendants héritent de cette direction, alors qu’ils n’ont toujours pas quitté le laboratoire, et l’hybridation entre deux populations ayant des orientations préférentielles différentes produit en F1 des descendants adoptant des orientations intermédiaires.Un travail de Scapini (1988) montre toutefois que, si les talitres présentent une composante d’orientation héritable, ils disposent aussi d’une grande plasticité épigénétique, qui varie, selon les individus testés, avec l’âge et l’apprentissage. Une canalisation progressive du comportement d’orientation, au cours du développement, par réduction de la variabilité intra-individuelle, conduit à une concentration des directions. Dans des conditions naturelles, ce processus semble plus strict et pertinent du point de vue écologique. Inversement, au laboratoire, après trente jours, les directions affichées par les individus diffèrent entre elles et ne correspondent pas nécessairement à la direction de la mer. La perte de l’orientation dominante dans les conditions d’élevage confirme l’importance d’une validation écologique dans la canalisation, à travers une interaction avec l’environnement. Toutefois, aucun de ces processus n’est irréversible, et il persiste encore des possibilités de modifier expérimentalement les choix bidirectionnels d’individus adultes déjà canalisés dans la direction de la mer familière. L’information génétiquement codée ne serait alors qu’une commande d’orientation dans la direction la plus probable du rivage pouvant interagir positivement avec l’apprentissage dans le déterminisme de l’adaptation. Les comportements optimaux, sélectionnés, peuvent ainsi résulter du couplage entre une large variabilité génétique et une grande plasticité individuelle.En étudiant l’orientation visuelle du grillon des bois dans son milieu naturel, Campan et al. (1987) ont montré que les déplacements, parfois répétitifs, selon des directions habituelles, étaient guidés, dans leur domaine vital, par des signaux individuellement appris: repères visuels terrestres, astronomiques et gravifiques. Ils ont pu valider des scénarios de développement dans lesquels l’orientation astronomique est construite sur la base d’une phototaxie négative initiale, l’utilisation des repères visuels terrestres à partir d’une scototaxie présente dès la naissance, et l’utilisation d’une direction de pente connue, à partir d’une géotaxie précoce. L’expression de ces tendances primitives, associées aux renforcements qu’elles procurent aux animaux, conduit chaque individu à construire, au cours de l’ontogenèse, son propre système de repères. Celui-ci devient évidemment dominant au sein d’une population habitant un lieu identique, même s’il persiste des différences interindividuelles et si un sujet peut modifier son expression d’un instant à l’autre.Chez d’autres espèces de grillons vivant sur les rives des lacs, un animal peut corriger à tout moment le repère astronomique qu’il utilise, selon l’orientation de la rive où il se trouve, d’où il saute à l’eau pour échapper aux dérangements et aux prédateurs (Beugnon, 1986). Le grillon champêtre, comme plusieurs espèces de fourmis, à chaque instant d’un trajet spontané hors de son terrier, quelle qu’en soit la complexité, est en mesure de revenir en ligne droite à son point de départ (Beugnon et Campan, 1989). Tout se passe comme s’il intégrait et mémorisait, au fur et à mesure de son déplacement, toutes les translations et les rotations du référentiel de son corps par rapport à un référentiel extérieur, qui est le plus souvent le plan de polarisation de la lumière solaire par le ciel.Toujours dans le domaine de l’ontogenèse de l’orientation des Insectes, les données classiques de la littérature révèlent des phénomènes plus riches encore. Les expériences fondatrices de Tinbergen (1932), complétées par un certain nombre d’autres sur les guêpes et les abeilles solitaires et sociales, démontrent que le retour au nid, donc sa localisation, est guidé par des configurations de repères proches que l’animal apprend pendant les quelques secondes que dure son envol; il fixe alors une image suffisamment précise de l’ensemble du panorama pour tolérer des modifications expérimentales de la situation, y compris la suppression des repères les plus proches, d’autres, plus lointains, assurant la suppléance.Épigenèse comportementaleLes processus mis en jeu dans l’ontogenèse comportementale sont au moins en partie homologues de ceux qui règlent le développement du système nerveux, mais, cette fois, avec une identification beaucoup plus explicite des facteurs environnementaux de l’épigenèse. On va retrouver des phénomènes (cf. EMBRYOLOGIE) sur le modèle de l’induction d’une étape du développement par la précédente, la détermination et la différenciation sélective de certains traits pendant des périodes sensibles, qui seront ensuite stabilisés ou non par la pratique et tous les renforcements associés, l’accélération ou le ralentissement des modifications comportementales, selon la richesse stimulante et renforçatrice de l’univers dans lequel l’organisme se développe. Ainsi, dans l’ontogenèse comportementale, aux processus de type embryologique vont se mêler tous ceux qui relèvent de l’apprentissage [cf. APPRENTISSAGE]: l’habituation et la sensibilisation, les conditionnements classiques et opérants, les apprentissages spatiaux et discriminatifs, les apprentissages à apprendre; s’ajoute à cela toute l’expérience individuelle acquise dans un cadre social [cf. SOCIÉTÉS ANIMALES], pour laquelle les parents (effets maternels ou paternels, au sens large, empreinte et attachement) ou d’autres congénères sont impliqués (observation, imitation, transmission «culturelle»), etc.Le développement comportemental va ainsi conduire à équiper progressivement chaque individu d’un certain nombre de programmes nerveux qui lui permettent d’abord d’assurer ses grandes fonctions biologiques. Mais la large plasticité dont fait preuve son système nerveux l’autorise, en même temps, à ajuster ses programmes aux conditions singulières de son histoire individuelle. La fonction biologique de cette plasticité est bien évidemment de lui permettre d’apprendre des relations causales liées aux objets et aux lieux, de résoudre des problèmes technologiques, mais également de saisir les nouveautés auxquelles il est fréquemment confronté afin d’y apporter des solutions immédiates qu’il mémorise.On peut appliquer à l’ontogenèse comportementale le même cadre théorique des processus auto-organisateurs que celui qui a été proposé précédemment pour la phylogenèse. Il consiste ici en une généralisation à l’ensemble des traits de comportement, d’un emboîtement de processus proche de celui proposé par Marler pour le chant des oiseaux, avec une structure primitive initiale commune à tous les individus de l’espèce («innée»), modifiée par un apprentissage lié aux conditions locales, donc comparable pour tous les individus d’une population («apprise»), puis par des acquis propres à chaque individu, qui le distinguent de ses voisins («inventée»), enfin par des modifications qu’apporte le sujet chaque fois qu’il réalise un comportement, qui tient compte des conditions immédiates particulières et uniques («improvisée»).Ainsi, à chaque instant, il est en mesure d’exprimer sa relation au monde par des manifestations comportementales fondées à la fois sur une activité nerveuse déjà préparée, mais que, par un traitement cognitif parfois complexe, il ajuste aux conditions qu’il rencontre au moment même de l’action, mais aussi à celles qu’il peut éventuellement prévoir du futur immédiat.4. Les causes immédiates du comportementLeur analyse répond à une autre des quatre questions posées par Tinbergen (1963).Les principes fondateursUne première formalisation est proposée par le schéma béhavioriste que nous avons déjà évoqué plus haut, où le stimulus est directement le déclencheur de la réponse, selon un certain nombre de lois dictées par la boîte noire.Dans le principe de la double quantification de Lorenz [cf. INSTINCT], un acte instinctif résulte de l’action de deux composantes, l’une, endogène, correspond à une énergie spécifique pour l’acte considéré, et l’autre, exogène, est le signal déclencheur responsable de la libération de cette énergie; les relations quantitatives entre ces deux composantes sont additives. L’hypothèse neuroéthologique associée suggère que l’énergie spécifique s’accumulerait en association avec un centre coordinateur inné de la commande pour l’exécution de l’acte. La perception du signal déclencheur correspondrait, plus ou moins exactement, à un modèle inné, représentation du signal naturel (Innate Releasing Pattern , I.R.P.), et activerait un mécanisme inné de déclenchement (I.R.M.) responsable de la libération de l’énergie spécifique et de l’exécution de l’acte sous une forme stéréotypée (F.A.P.).Tinbergen, qui généralise ce principe à l’ensemble des comportements instinctifs, propose une organisation nerveuse hiérarchique des commandes comportementales, des I.R.M. et des I.R.P., les niveaux supérieurs étant liés par des interactions inhibitrices et les inférieurs coordonnant les séquences, l’exécution d’un acte étant le déclencheur du suivant [cf. INSTINCT].Élargissement des concepts fondateursLe concept de stimulus signal, déclencheur du comportement, est certainement le plus fécond de tous ceux issus de l’objectivisme, et il a réussi à franchir les décennies sans prendre trop de rides. Il est souvent nommé déclencheur (releaser ) et représente un stimulus ou une configuration perceptive (image de recherche) de l’Umwelt d’un animal, capable d’évoquer un acte moteur ou une séquence comportementale particulière. Ce concept a généré sa propre méthode expérimentale, la méthode des leurres. Le succès du concept comme de la méthode s’explique parce qu’ils représentent un moyen unique d’explorer l’univers propre des êtres sans langage.Toutefois, si la méthode est toujours actuelle, le concept a été sensiblement nuancé par rapport à la vision initiale. Le déclencheur n’est plus ce modèle simple, grossier, capable d’évoquer, d’une manière presque réflexe, la réponse comportementale. Le fait qu’il soit efficace ne signifie plus que l’animal le confonde avec l’objet naturel et toutes ses caractéristiques. De plus, le stimulus signal n’est plus isolé de son contexte, et sa reconnaissance peut dépendre très largement de sa position et de ses relations figurales avec toute la constellation de stimulations qui sollicite en permanence le sujet. Enfin, il n’est plus seulement considéré comme spécifique de l’espèce, inné et héritable, mais aussi construit au cours du développement individuel, sous la forme d’une représentation à tout instant révisable par apprentissage ou généralisation perceptive.Ewert (1984) rapporte l’analyse neuroéthologique de l’I.R.M. pour le déclenchement de la capture chez le crapaud, montrant que les détecteurs de traits sont organisés sous la forme de réseaux limités de neurones interconnectés (modules), entre le toit optique (T) et le thalamus (TH). Il a pu établir des séquences cellulaires pour divers aspects du comportement:
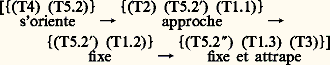 ou bien,
ou bien,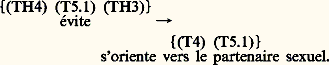 Par ailleurs, il a complété le concept d’I.R.M. par celui d’I.R.Me, modifié par l’expérience, et celui d’A.R.M., mécanisme acquis de déclenchement.Holley et Gervais (1986) ont recherché les traces structurales et fonctionnelles d’une image de recherche apprise, dans l’épithélium olfactif du rat, à la sortie de la couche glomérulaire. Il s’agit d’une représentation de l’odeur du groupe dont la fonction, dans le comportement social, est démontrée. Aucun stimulus ne dispose d’un canal spécifique de transmission, mais la mesure de la densité neuronale et de sa distribution spatiale, dans le neuroépithélium olfactif et à la sortie du bulbe, permet de construire une image caractéristique (fig. 8).Le concept de F.A.P. n’est relativement fixe que pour des actes moteurs élémentaires à fonction générale (marche, saut, etc.) ou bien pour des signaux produits par un émetteur dans un comportement de communication où il doit être décodé par un récepteur [cf. COMMUNICATION]. Aussi, Barlow (1968) a proposé de lui substituer celui de M.A.P. patron d’action modal, qui indique la parenté des structures comportementales pour tous les individus d’une même espèce, mais laisse ouvertes toutes les possibilités de différences inter- et intra-individuelles. Enfin, dès qu’il s’agit d’une séquence comportementale enchaînant plusieurs actes, l’organisation et la coordination en sont souvent variables et modifiables par apprentissage.Durant l’exécution de l’acte, le mouvement peut être contrôlé et ajusté sur la base de réafférences musculaires immédiates qui sont comparées à des copies d’efférence, c’est-à-dire de l’ordre moteur (von Holst et Mittelstaedt, 1953). Tout écart entre réafférence et copie d’efférence donne lieu à une correction de la commande [cf. MOTRICITÉ]. Enfin, les effets en retour des conséquences de l’acte vont conduire soit à la poursuite, soit à l’arrêt du comportement engagé.En ce qui concerne l’énergie spécifique de la double quantification, elle a donné lieu, dans ses relations avec les déclencheurs, à l’élaboration de divers modèles de motivation.Un premier examen du concept a été fait par Baerends et al. (1955) lors d’une étude de la parade nuptiale du poisson guppy. La condition reproductrice du mâle est évaluée par sa coloration, qui peut changer rapidement (composante endogène). La femelle est d’autant plus attractive qu’elle est grosse (composante exogène). Les mâles courtisent les femelles en utilisant tout un répertoire de signaux gradués dont le niveau traduit l’additivité des deux composantes (fig. 9).Avec l’augmentation de sa motivation sexuelle, le mâle répond à des femelles plus petites, même si les plus grandes restent les plus efficaces pour déclencher: les mouvements sigmoïdes complets (S), les mouvements sigmoïdes d’intention (Si) ou le placement (P). Un mâle faiblement motivé a besoin d’un stimulus extérieur plus évocateur qu’un mâle fortement motivé, ce qui correspond parfaitement au modèle de la double quantification. Les courbes qui relient les points correspondant à un même niveau de tendance, c’est-à-dire à l’ensemble des combinaisons entre causes externes et facteurs internes donnant une même valeur résultante, sont décrites comme des isoclines motivationnelles (McFarland, 1976).Le prolongement le plus riche de la double quantification se retrouve dans les modèles proposés par les conflits de motivations. On admet généralement que l’animal ne peut réaliser qu’une activité à la fois. Lorsque deux tendances sont en jeu simultanément, elles génèrent un conflit, qui peut être de type «approche-approche», «évitement-évitement» ou «approche-évitement». La décision comportementale finale de l’animal correspond soit à l’une des tendances, soit aux deux successivement, soit, enfin, à des comportements différents, inadéquats par rapport à la situation (activités substitutives ou redirigées: displacement activities ).McFarland et Sibly (1975) ont proposé le modèle d’espaces d’états (state space approach ) pour expliquer la solution des conflits de motivation (fig. 10). Il permet, sur la base d’informations quantitatives simples, de prévoir la décision de l’animal; Pring-Mill (1979) l’a généralisé à tout changement d’activité en introduisant des possibilités d’apprentissage.Quel que soit leur niveau d’élaboration, les modèles précédents représentent d’une manière caricaturale les processus immédiats qui conduisent un animal à réaliser un comportement. Des quantités d’autres variables intermédiaires, se modifiant avec le temps, participent à la dimension motivationnelle du déclenchement comportemental: niveau d’activation et rythmes d’activité, état endocrinien [cf. SEXUEL (COMPORTEMENT)], émotivité, etc., auxquels s’ajoutent toutes les influences de l’expérience individuelle et de la sphère cognitive.L’organisation centrale des comportements en termes de niveaux hiérarchiques pré-organisés, telle que l’avait proposée Tinbergen, doit également être revue.Si elle pouvait convenir à des séquences stéréotypées, la construction des diagrammes de déroulement (Baerends et al., 1955, et bien d’autres après lui) ainsi que l’évaluation quantitative des fréquences de transition entre les actes d’une séquence en montrent la très grande variabilité d’organisation.Toutefois, chaque acte reste identifiable (M.A.P.) et correspond à une structure, relativement constante liée à un module de commande. Cela peut être illustré par les travaux sur le chant des oiseaux ou celui des grillons (Huber), ou, bien sûr, des manifestations motrices simples, comme la gloutonnerie chez le pleurobranche (Gillette, 1987). On peut, sans doute, défendre aujourd’hui l’idée qu’une organisation nerveuse modulaire, à des niveaux relativement périphériques, à la fois structurale et fonctionnelle sous-tend l’exécution des unités comportementales composant les séquences. Distinguer des niveaux périphériques et d’autres plus centraux implique encore une certaine hiérarchisation. Mais la mise en jeu des modules, dans un ordre souvent indéterminé, à n’importe quel moment, révèle toute une plasticité que seul un système en réseau peu hiérarchisé peut exprimer. Par ailleurs, des données neurophysiologiques apportent des arguments qui nuancent la vision hiérarchique ainsi que le concept même de module. Par exemple, le retrait du manteau chez l’aplysie, dont on pensait jusqu’ici qu’il mettait en jeu seulement quelques neurones bien localisés, implique en fait au moins cent cinquante unités, comme le montrent les méthodes d’enregistrement optique. De même l’imagerie cérébrale chez les Primates, lors de l’activité mentale ou motrice d’un sujet, si elle fait apparaître une dominante dans certaines zones, montre que d’autres régions sont activées et qu’il existe une importante variabilité inter- et intra-individuelle.Enfin, l’animal réalise des opérations mentales de traitement des informations perçues et représentées, avant ou au moment de décider d’une action motrice. On doit rappeler les anciennes expériences de W. Koehler sur les singes attrapant un objet à l’aide d’outils, de O. Koehler sur l’apprentissage à compter chez les Oiseaux, de Lashley sur la généralisation perceptive chez les rats ou de Tolman sur les cartes mentales. Plus récemment, l’utilisation d’un raccourci a été démontrée chez le chien par Fabrigoule (1985). Lors d’une série d’expériences préparatoires, les chiens, tenus en laisse, sont entraînés à retrouver de la nourriture, disposée dans des coupelles, selon une séquence déterminée (fig. 11), par exemple en A, en B, puis en D. Lorsque l’apprentissage est terminé, les animaux sont lâchés au centre (O) et laissés libres de développer leur propre stratégie spatiale; chaque branche du dispositif en étoile contient un plat, mais seuls A, B et D sont renforcés, selon la séquence apprise la veille. Si le chien commence par A, il gagnera ensuite B, puis D en utilisant un raccourci, trajet inventé qu’il n’a jamais parcouru avec l’expérimentateur.Une autre illustration du traitement mental des représentations peut être tirée d’une expérience de transfert intermodalitaire entre la vision et le toucher chez les Primates, rapportée par Vauclair (1987) d’après les travaux de Davenport.La technique utilisée est celle de l’appariement à un modèle qui permet une comparaison immédiate ou différée des stimulus dans les deux modalités. Trois objets, par exemple, sont présentés au sujet: il peut en voir deux et toucher le troisième; l’un des objets visuels est identique à celui qui est exploré par voie tactilo-kinesthésique, tandis que l’autre est nettement différent. La tâche consiste à toucher un objet, à observer les deux autres et, finalement, à les apparier en allongeant la main dans la bonne direction. Si le choix est correct, le singe reçoit une récompense alimentaire. On réalise ainsi un pré-apprentissage pour l’exploration tactile, l’observation visuelle et la réponse de choix. Ensuite, on procède à un grand nombre d’essais avec l’ensemble des trois objets, en présentant de façon aléatoire celui à apparier. Enfin, les singes sont soumis à un minimum de quarante problèmes uniques (un seul essai) en recourant chaque fois à des objets nouveaux. Chaque expérience peut naturellement être abordée de façon bidirectionnelle commençant par l’exploration visuelle ou tactile. Quatre résultats méritent d’être rapportés: 1. les animaux sont capables de perception intermodale pour des objets tridimensionnels, quelle que soit la modalité avec laquelle on effectue le test; 2. cette capacité à reconnaître l’équivalence entre objets perçus à travers des canaux sensoriels différents se maintient lorsque le test est réalisé avec des photographies grandeur nature qui les représentent; 3. si l’on dégrade certaines propriétés du stimulus en utilisant, par exemple, des photographies très contrastées, des photos demi-format ou encore une simple esquisse de l’objet, les singes réussissent le transfert; 4. l’introduction d’un délai de quinze à trente secondes entre la présentation dans une modalité et la reconnaissance dans l’autre n’altère pas significativement les performances.La capacité de l’animal à établir l’identification d’un objet à partir d’informations provenant de deux canaux sensoriels distincts, et surtout à généraliser cette équivalence sur la base de propriétés limitées des objets, suggère que le singe a bien constitué une sorte de représentation mentale, un «concept d’un stimulus».Toute cette sphère cognitive chez l’animal et la plasticité des réseaux nerveux sous-jacents sont aujourd’hui des chapitres importants de l’étude du comportement, qui se développent en étroite liaison avec les sciences plus formelles comme l’intelligence artificielle, l’analyse d’image, etc. Il y a beaucoup à attendre de cette démarche.La manifestation comportementale hic et nunc et son cortège de causes constituent un maillon essentiel dans l’auto-organisation des programmes nerveux qui la sous-tendent. Donc, non seulement ils s’inscrivent dans la dynamique à la fois phylogénétique et ontogénétique, mais participent également à leur tour à ces processus, en contribuant à la construction des phénotypes individuels, puis à l’assimilation génétique. L’étude du comportement a donc répondu, au moins en partie, aux quatre questions posées par Tinbergen en 1963 qui en délimitaient l’objet.
Par ailleurs, il a complété le concept d’I.R.M. par celui d’I.R.Me, modifié par l’expérience, et celui d’A.R.M., mécanisme acquis de déclenchement.Holley et Gervais (1986) ont recherché les traces structurales et fonctionnelles d’une image de recherche apprise, dans l’épithélium olfactif du rat, à la sortie de la couche glomérulaire. Il s’agit d’une représentation de l’odeur du groupe dont la fonction, dans le comportement social, est démontrée. Aucun stimulus ne dispose d’un canal spécifique de transmission, mais la mesure de la densité neuronale et de sa distribution spatiale, dans le neuroépithélium olfactif et à la sortie du bulbe, permet de construire une image caractéristique (fig. 8).Le concept de F.A.P. n’est relativement fixe que pour des actes moteurs élémentaires à fonction générale (marche, saut, etc.) ou bien pour des signaux produits par un émetteur dans un comportement de communication où il doit être décodé par un récepteur [cf. COMMUNICATION]. Aussi, Barlow (1968) a proposé de lui substituer celui de M.A.P. patron d’action modal, qui indique la parenté des structures comportementales pour tous les individus d’une même espèce, mais laisse ouvertes toutes les possibilités de différences inter- et intra-individuelles. Enfin, dès qu’il s’agit d’une séquence comportementale enchaînant plusieurs actes, l’organisation et la coordination en sont souvent variables et modifiables par apprentissage.Durant l’exécution de l’acte, le mouvement peut être contrôlé et ajusté sur la base de réafférences musculaires immédiates qui sont comparées à des copies d’efférence, c’est-à-dire de l’ordre moteur (von Holst et Mittelstaedt, 1953). Tout écart entre réafférence et copie d’efférence donne lieu à une correction de la commande [cf. MOTRICITÉ]. Enfin, les effets en retour des conséquences de l’acte vont conduire soit à la poursuite, soit à l’arrêt du comportement engagé.En ce qui concerne l’énergie spécifique de la double quantification, elle a donné lieu, dans ses relations avec les déclencheurs, à l’élaboration de divers modèles de motivation.Un premier examen du concept a été fait par Baerends et al. (1955) lors d’une étude de la parade nuptiale du poisson guppy. La condition reproductrice du mâle est évaluée par sa coloration, qui peut changer rapidement (composante endogène). La femelle est d’autant plus attractive qu’elle est grosse (composante exogène). Les mâles courtisent les femelles en utilisant tout un répertoire de signaux gradués dont le niveau traduit l’additivité des deux composantes (fig. 9).Avec l’augmentation de sa motivation sexuelle, le mâle répond à des femelles plus petites, même si les plus grandes restent les plus efficaces pour déclencher: les mouvements sigmoïdes complets (S), les mouvements sigmoïdes d’intention (Si) ou le placement (P). Un mâle faiblement motivé a besoin d’un stimulus extérieur plus évocateur qu’un mâle fortement motivé, ce qui correspond parfaitement au modèle de la double quantification. Les courbes qui relient les points correspondant à un même niveau de tendance, c’est-à-dire à l’ensemble des combinaisons entre causes externes et facteurs internes donnant une même valeur résultante, sont décrites comme des isoclines motivationnelles (McFarland, 1976).Le prolongement le plus riche de la double quantification se retrouve dans les modèles proposés par les conflits de motivations. On admet généralement que l’animal ne peut réaliser qu’une activité à la fois. Lorsque deux tendances sont en jeu simultanément, elles génèrent un conflit, qui peut être de type «approche-approche», «évitement-évitement» ou «approche-évitement». La décision comportementale finale de l’animal correspond soit à l’une des tendances, soit aux deux successivement, soit, enfin, à des comportements différents, inadéquats par rapport à la situation (activités substitutives ou redirigées: displacement activities ).McFarland et Sibly (1975) ont proposé le modèle d’espaces d’états (state space approach ) pour expliquer la solution des conflits de motivation (fig. 10). Il permet, sur la base d’informations quantitatives simples, de prévoir la décision de l’animal; Pring-Mill (1979) l’a généralisé à tout changement d’activité en introduisant des possibilités d’apprentissage.Quel que soit leur niveau d’élaboration, les modèles précédents représentent d’une manière caricaturale les processus immédiats qui conduisent un animal à réaliser un comportement. Des quantités d’autres variables intermédiaires, se modifiant avec le temps, participent à la dimension motivationnelle du déclenchement comportemental: niveau d’activation et rythmes d’activité, état endocrinien [cf. SEXUEL (COMPORTEMENT)], émotivité, etc., auxquels s’ajoutent toutes les influences de l’expérience individuelle et de la sphère cognitive.L’organisation centrale des comportements en termes de niveaux hiérarchiques pré-organisés, telle que l’avait proposée Tinbergen, doit également être revue.Si elle pouvait convenir à des séquences stéréotypées, la construction des diagrammes de déroulement (Baerends et al., 1955, et bien d’autres après lui) ainsi que l’évaluation quantitative des fréquences de transition entre les actes d’une séquence en montrent la très grande variabilité d’organisation.Toutefois, chaque acte reste identifiable (M.A.P.) et correspond à une structure, relativement constante liée à un module de commande. Cela peut être illustré par les travaux sur le chant des oiseaux ou celui des grillons (Huber), ou, bien sûr, des manifestations motrices simples, comme la gloutonnerie chez le pleurobranche (Gillette, 1987). On peut, sans doute, défendre aujourd’hui l’idée qu’une organisation nerveuse modulaire, à des niveaux relativement périphériques, à la fois structurale et fonctionnelle sous-tend l’exécution des unités comportementales composant les séquences. Distinguer des niveaux périphériques et d’autres plus centraux implique encore une certaine hiérarchisation. Mais la mise en jeu des modules, dans un ordre souvent indéterminé, à n’importe quel moment, révèle toute une plasticité que seul un système en réseau peu hiérarchisé peut exprimer. Par ailleurs, des données neurophysiologiques apportent des arguments qui nuancent la vision hiérarchique ainsi que le concept même de module. Par exemple, le retrait du manteau chez l’aplysie, dont on pensait jusqu’ici qu’il mettait en jeu seulement quelques neurones bien localisés, implique en fait au moins cent cinquante unités, comme le montrent les méthodes d’enregistrement optique. De même l’imagerie cérébrale chez les Primates, lors de l’activité mentale ou motrice d’un sujet, si elle fait apparaître une dominante dans certaines zones, montre que d’autres régions sont activées et qu’il existe une importante variabilité inter- et intra-individuelle.Enfin, l’animal réalise des opérations mentales de traitement des informations perçues et représentées, avant ou au moment de décider d’une action motrice. On doit rappeler les anciennes expériences de W. Koehler sur les singes attrapant un objet à l’aide d’outils, de O. Koehler sur l’apprentissage à compter chez les Oiseaux, de Lashley sur la généralisation perceptive chez les rats ou de Tolman sur les cartes mentales. Plus récemment, l’utilisation d’un raccourci a été démontrée chez le chien par Fabrigoule (1985). Lors d’une série d’expériences préparatoires, les chiens, tenus en laisse, sont entraînés à retrouver de la nourriture, disposée dans des coupelles, selon une séquence déterminée (fig. 11), par exemple en A, en B, puis en D. Lorsque l’apprentissage est terminé, les animaux sont lâchés au centre (O) et laissés libres de développer leur propre stratégie spatiale; chaque branche du dispositif en étoile contient un plat, mais seuls A, B et D sont renforcés, selon la séquence apprise la veille. Si le chien commence par A, il gagnera ensuite B, puis D en utilisant un raccourci, trajet inventé qu’il n’a jamais parcouru avec l’expérimentateur.Une autre illustration du traitement mental des représentations peut être tirée d’une expérience de transfert intermodalitaire entre la vision et le toucher chez les Primates, rapportée par Vauclair (1987) d’après les travaux de Davenport.La technique utilisée est celle de l’appariement à un modèle qui permet une comparaison immédiate ou différée des stimulus dans les deux modalités. Trois objets, par exemple, sont présentés au sujet: il peut en voir deux et toucher le troisième; l’un des objets visuels est identique à celui qui est exploré par voie tactilo-kinesthésique, tandis que l’autre est nettement différent. La tâche consiste à toucher un objet, à observer les deux autres et, finalement, à les apparier en allongeant la main dans la bonne direction. Si le choix est correct, le singe reçoit une récompense alimentaire. On réalise ainsi un pré-apprentissage pour l’exploration tactile, l’observation visuelle et la réponse de choix. Ensuite, on procède à un grand nombre d’essais avec l’ensemble des trois objets, en présentant de façon aléatoire celui à apparier. Enfin, les singes sont soumis à un minimum de quarante problèmes uniques (un seul essai) en recourant chaque fois à des objets nouveaux. Chaque expérience peut naturellement être abordée de façon bidirectionnelle commençant par l’exploration visuelle ou tactile. Quatre résultats méritent d’être rapportés: 1. les animaux sont capables de perception intermodale pour des objets tridimensionnels, quelle que soit la modalité avec laquelle on effectue le test; 2. cette capacité à reconnaître l’équivalence entre objets perçus à travers des canaux sensoriels différents se maintient lorsque le test est réalisé avec des photographies grandeur nature qui les représentent; 3. si l’on dégrade certaines propriétés du stimulus en utilisant, par exemple, des photographies très contrastées, des photos demi-format ou encore une simple esquisse de l’objet, les singes réussissent le transfert; 4. l’introduction d’un délai de quinze à trente secondes entre la présentation dans une modalité et la reconnaissance dans l’autre n’altère pas significativement les performances.La capacité de l’animal à établir l’identification d’un objet à partir d’informations provenant de deux canaux sensoriels distincts, et surtout à généraliser cette équivalence sur la base de propriétés limitées des objets, suggère que le singe a bien constitué une sorte de représentation mentale, un «concept d’un stimulus».Toute cette sphère cognitive chez l’animal et la plasticité des réseaux nerveux sous-jacents sont aujourd’hui des chapitres importants de l’étude du comportement, qui se développent en étroite liaison avec les sciences plus formelles comme l’intelligence artificielle, l’analyse d’image, etc. Il y a beaucoup à attendre de cette démarche.La manifestation comportementale hic et nunc et son cortège de causes constituent un maillon essentiel dans l’auto-organisation des programmes nerveux qui la sous-tendent. Donc, non seulement ils s’inscrivent dans la dynamique à la fois phylogénétique et ontogénétique, mais participent également à leur tour à ces processus, en contribuant à la construction des phénotypes individuels, puis à l’assimilation génétique. L’étude du comportement a donc répondu, au moins en partie, aux quatre questions posées par Tinbergen en 1963 qui en délimitaient l’objet.
Encyclopédie Universelle. 2012.